Č«ĘŔ˝çÔĽÓĐ30-40%µÄżÉ¸űµŘşÍ50%µÄDZÔڿɸűµŘĘôÓÚËáĐÔÍÁČŔ[1]Ł¬ÂÁŔë×Ó¶ľş¦ĘǸĂŔŕÍÁČŔÖĐĎŢÖĆ×÷ÎďÉú˛úµÄÖ÷ŇŞŇňËŘ[2]ˇŁÔÚł¤ĆÚĘĘÓ¦ąýłĚÖĐŁ¬Ö˛Îď˝ř»ŻłöÁ˶ŕÖÖÄÍÂÁ»úÖơŁĆäÖĐŁ¬Ö˛Îďͨąý¸ůĽâϸ°űĎňÍâ·ÖĂÚĆ»ąűËáµČÓĐ»úËáňüşĎ¸ůĽĘÍÁČŔČÜŇşÖеÄÂÁŔë×ÓŁ¬´Ó¶řĎűłýĆ䶾ĐÔŁ¬ĘÇÖ˛ÎďµÖÓůÂÁ¶ľµÄÖ÷ŇŞÍľľ¶[2,3]ˇŁÔçÔÚ2004ÄęŃĐľżŐ߾ͷ˘ĎÖÖ˛Îď¸ůϸ°űÖĐÂÁĽ¤»îµÄĆ»ąűËá·ÖĂÚĘÇÓÉĆ»ąűËáתÔ˵°°×Ł¨Aluminum-activated Malate TransporterŁ¬ALMT1Ł©˝éµĽ[4]ˇŁÖ®şóŁ¬ŃĐľżŐß֤ʵÁËÔÚСÂóˇ˘ÄâÄϽ桢ÓͲˡ˘´ó¶ąˇ˘şÚÂóµČ¶ŕÖÖÖ˛ÎďÖиûúÖĆľßÓбŁĘŘĐÔˇŁµ«ĘÇŁ¬ł¤ĆÚŇÔŔ´ČËĂǶÔÓÚÂÁŔë×ÓČçşÎĽ¤»îALMT1תÔËĆ»ąűËáČÔ˛»µĂ¶řÖŞˇŁ
2021Äę11ÔÂ19ČŐŁ¬Őă˝´óѧąů˝ĚΡ˘ŃîΡşÍÖŁÉÜ˝¨Čý¸öżÎĚâ×é×éłÉµÄşĎ×÷ÍŶÓÔÚCell ResearchÉĎÔÚĎß·˘±íÁËĚâÎŞStructural Basis of ALMT1-Mediated Aluminum Resistance in ArabidopsisµÄŃĐľżÂŰÎÄŁ¬˝âÎöÁËÄâÄĎ˝ćALMT1ͨµŔµ°°×Ł¨AtALMT1Ł©¶ŕÖÖ״̬ϵÄČýά˝áąąŁ¬˝áşĎµçÉúŔíʵŃ顢·Ö×Ó¶ŻÁ¦Ń§ÄŁÄâşÍÄâÄĎ˝ćĚĺÄÚʵŃ飬˛űĂ÷ÁËAtALMT1˝éµĽµÄÂÁĽ¤»îĆ»ąűËáתÔ˵ķÖ×Ó»úŔíˇŁ
¸ĂŃĐľż±íĂ÷Ł¬AtALMT1˝áąąĘÇŇ»¸öĐÂĐ͵ÄŇőŔë×ÓͨµŔ˝áąąˇŁAtALMT1ŇÔ¶ţľŰĚĺĐÎĘ˝×é×°łÉŇ»¸öŇőŔë×ÓͨµŔŁ¬Ăż¸öŃÇ»ů°üş¬6¸öżçĤÂÝĐýŁ¨TM1-6Ł©şÍ6¸öϸ°űÖʲŕµÄÂÝĐýŁ¨H1-H6Ł©ˇŁŔ´×ÔÁ˝¸öŃÇ»ůµÄĘ®¶ţ¸öżçĤÂÝĐý×é×°łÉŔë×Óͨ͸ż×µŔŁ¨ÍĽ1Ł©ˇŁ

ÍĽ1 AtALMT1µÄ˝áąąˇŁAŁ¬AtALMT1µÄŔ䶳µçľµČýάÖŘąąŁ¨×󣩺Ϳ¨Í¨ÄŁĐÍŁ¨ÓŇŁ©µÄ˛ŕĘÓÍĽŁ¨ÉĎŁ©şÍ¸©ĘÓÍĽŁ¨ĎÂŁ©ˇŁBŁ¬AtALMT1µĄ¸öŃÇ»ůµÄ˛ŕĘÓÍĽˇŁCŁ¬AtALMT1µÄÍŘĆ˽ṹˇŁ
ÔÚĆ»ąűËá˝áşĎµÄAtALMT1˝áąąÖĐŁ¬ŃĐľżČËÔ±ąŰ˛ěµ˝Ć»ąűËáλÓÚAtALMT1µÄŔë×Óͨ͸ż×µŔÖм䣬ÓëÁ˝¶Ôľ«°±Ëá˛Đ»ůŁ¨Arg80şÍArg165Ł©Ď໥×÷ÓĂŁ¨ÍĽ2AŁ¬2BŁ©ˇŁŃĐľżČËԱͨąý·Ö×Ó¶ŻÁ¦Ń§ÄŁÄâşÍµçÉúŔíʵŃéŃéÖ¤ÁËŐâһƻąűËá˝áşĎλµăˇŁÔÚµçÉúŔíʵŃéÖĐŁ¬ŐâÁ˝¸öąŘĽüľ«°±ËáµÄÍ»±ä¶ĽÄÜĎÔÖřÓ°ĎěAtALMT1¶ÔĆ»ąűËáµÄͨ͸Ł¬ĆäÖĐArg165Í»±äĘąAtALMT1ÍęȫɥʧƻąűËá´«µĽµÄą¦ÄÜŁ¨ÍĽ2CŁ©ˇŁ
ÔÚÂÁŔë×Ó˝áşĎµÄAtALMT1˝áąąµ±ÖĐŁ¬ŃĐľżČËÔ±·˘ĎÖÂÁŔë×ÓÖ÷ŇŞ˝áşĎÔÚAtALMT1µÄϸ°űÍâ˛ŕŁ¬ÓÉTM1-2 loopşÍTM6ÉϵÄČý¸ö´ř¸şµçµÄ°±»ůËá˛Đ»ůĹäλŁ¨Asp40ˇ˘Glu156şÍAsp160Ł©Ł¨ÍĽ2DŁ¬2EŁ©ˇŁµçÉúŔíʵŃé±íĂ÷Ł¬ŐâČý¸öλµă°±»ůËáµÄÍ»±ä»áĎÔÖř˝µµÍAtALMT1˝éµĽµÄÂÁĽ¤»îµÄĆ»ąűËáµÄͨ͸Ł¨ÍĽ2Ł©ˇŁ
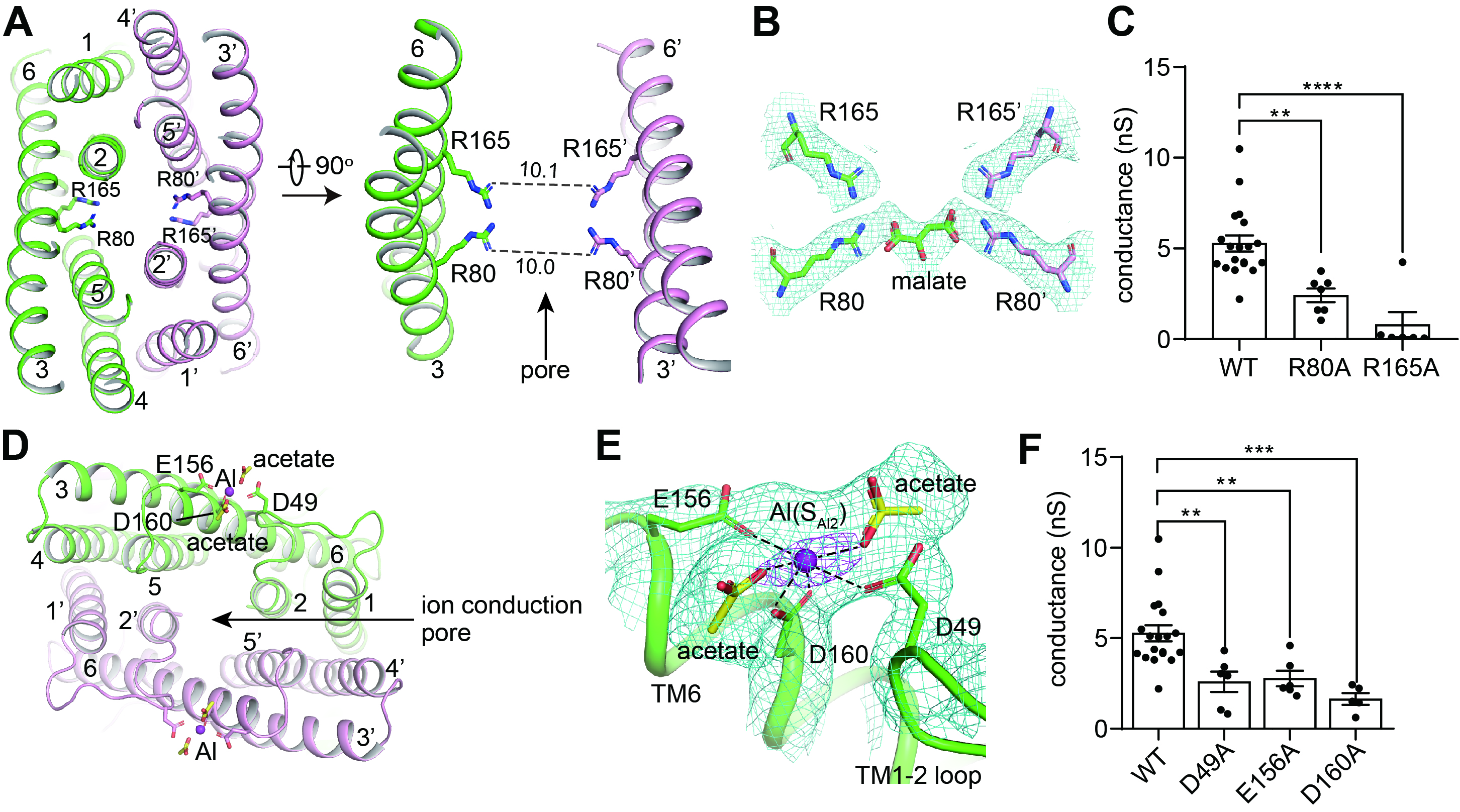
ÍĽ2 AtALMT1ÖĐĆ»ąűËáşÍϸ°űÍâ˛ŕÂÁŔë×ӵĽáşĎλµăˇŁAŁ¬Ŕë×Óͨ͸ż×µŔÖеÄŇ»¶Ô±ŁĘصľ«°±ËáArg80şÍArg165ˇŁBŁ¬Ć»ąűËáÓëÁ˝¶Ôľ«°±ËáĎ໥×÷ÓáŁCŁ¬ľ«°±ËáArg80şÍArg165Í»±äĎÔÖř˝µµÍAtALMT1¶ÔĆ»ąűËáµÄͨ͸ˇŁDŁ¬AtALMT1ÖĐϸ°űÍâ˛ŕÂÁŔë×ӵĽáşĎλµăˇŁEŁ¬ÂÁŔë×ÓÓëÖÜΧµÄËáĐÔ°±»ůËáAsp40ˇ˘Glu156şÍAsp160Ď໥×÷ÓáŁFŁ¬Asp40ˇ˘Glu156şÍAsp160Í»±äĎÔÖř˝µµÍAtALMT1˝éµĽµÄÂÁĽ¤»îµÄĆ»ąűËáµÄͨ͸ˇŁ
ÎŞŃéÖ¤AtALMT1ÖĐĆ»ąűËáşÍÂÁŔë×Ó˝áşĎλµăµÄ°±»ůËáµÄ×÷ÓĂŁ¬ŃĐľżČËÔ±ĹŕÓýÁËŐâĐ©°±»ůËáÍ»±äµÄÄâÄĎ˝ćת»ůŇňÖęϵˇŁ·˘ĎÖÔÚĂ»ÓĐÂÁŔë×ÓµÄÇéżöĎÂŁ¬Í»±äϵ±íĎÖłöÓëŇ°ÉúĐÍĎŕËƵĸůÉúł¤şÍĆ»ąűËá·ÖĂÚŁ»ÔÚÂÁŔë×Ó´¦ŔíşóŁ¬Í»±äϵ±íĎÖłöĎÔÖř˝µµÍµÄ¸ůÉúł¤şÍĆ»ąűËá·ÖĂÚŁ¬±íĎÖłöÓ빦ÄÜȱʧͻ±äĚĺalmt1ĎŕËƵıíĐÍŁ¨ÍĽ3Ł©ˇŁŇň´ËŁ¬ŐâĐ©ĚĺÄÚʵŃé˝řŇ»˛˝Č·ČĎÁËĆ»ąűËáşÍÂÁŔë×Ó˝áşĎλµăµÄ°±»ůËáÔÚÂÁĽ¤»îĆ»ąűËáתÔËÖеÄÖŘŇŞ×÷ÓáŁ
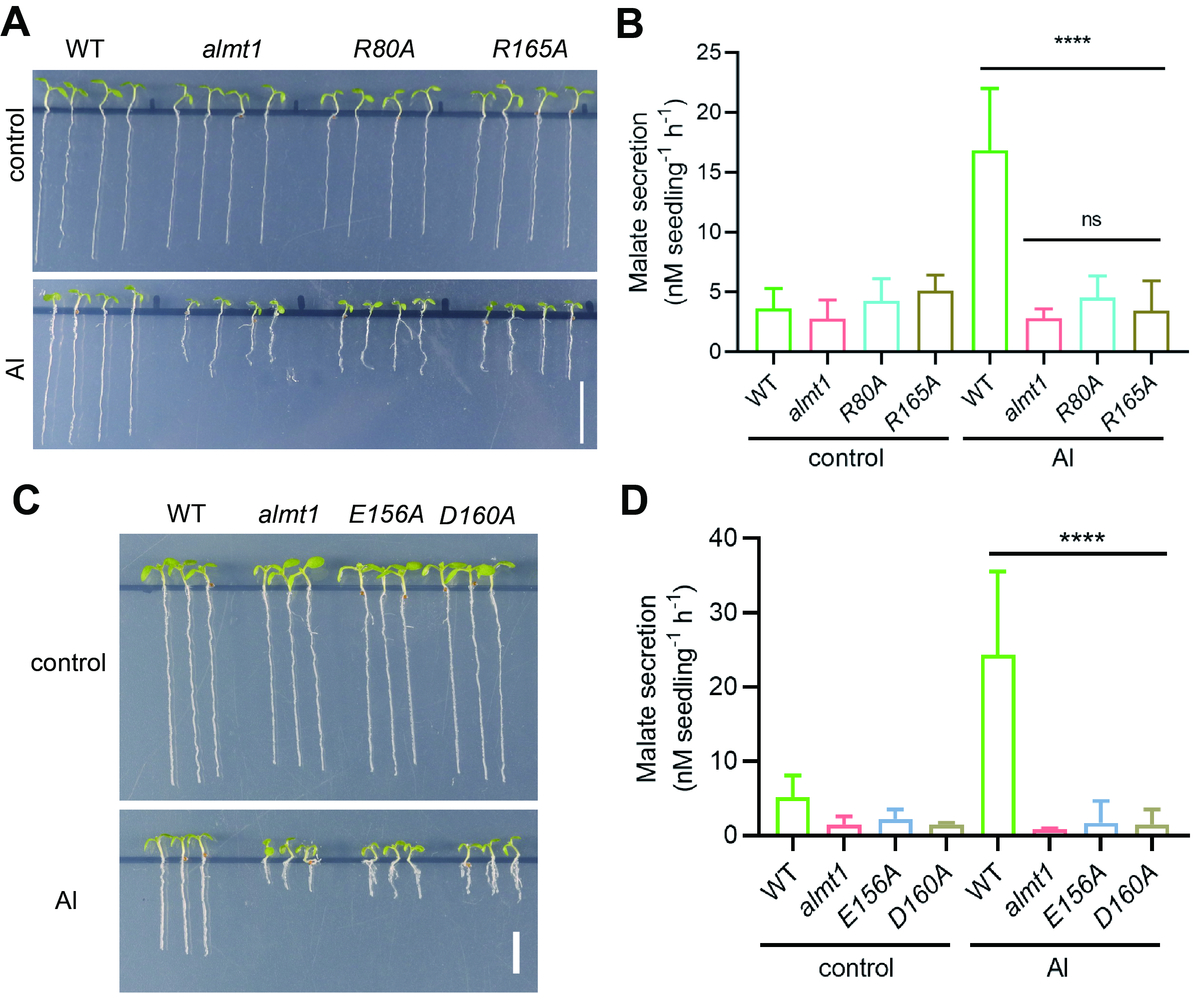
ÍĽ3 ÄâÄĎ˝ćĚĺÄÚŃéÖ¤AtALMT1ÖĐĆ»ąűËáşÍÂÁŔë×Ó˝áşĎλµăµÄ°±»ůËáµÄ×÷ÓáŁA-BŁ¬Ň°ÉúĐ͡˘AtALMT1ȱʧͻ±äÖęalmt1ˇ˘Ć»ąűËá˝áşĎλµăľ«°±ËáÍ»±äÖęR80AşÍR165AµÄ¸ůϵÉúł¤Ł¨AŁ©şÍĆ»ąűËá·ÖĂÚŁ¨BŁ©ˇŁC-DŁ¬Ň°ÉúĐ͡˘AtALMT1ȱʧͻ±äÖęalmt1ˇ˘ÂÁŔë×Ó˝áşĎλµăÍ»±äÖęE165AşÍD160AµÄ¸ůϵÉúł¤Ł¨CŁ©şÍĆ»ąűËá·ÖĂÚŁ¨DŁ©ˇŁ
δ˝áşĎÂÁŔë×ÓµÄAtALMT1´¦ÓڹرŐ״̬Ł¬˝áşĎÂÁŔë×ÓµÄAtALMT1ͨµŔ´¦ÓÚżŞ·Ĺ״̬ˇŁÍ¨ąý±Č˝ĎąŘ±Ő״̬şÍżŞ·Ĺ״̬µÄAtALMT1µÄ˝áąąŁ¬ŃĐľżČËÔ±ĚáłöÁËÂÁŔë×ÓĽ¤»îAtALMT1µÄ·Ö×Ó»úÖơŁÔÚËáĐÔĚőĽţĎÂŁ¬ÂÁŔë×Ó˝áşĎÔÚAtALMT1µÄ°űÍâ˛ŕTM1-2 loopşÍTM6Ö®ĽäŁ¬ÓëČý¸öËáĐԲлůŁ¨Asp40ˇ˘Glu156şÍAsp160Ł©ĹäλŁ¬ŇýĆđTM1-2 loopşÍTM5-6 loopąąĎó±ä»ŻŁ¬´Ó¶řµĽÖÂIle53˛Đ»ůĎňÍâÔ˶ŻŁ¬ŇýĆđż×µŔżŞ·ĹŁ¨ÍĽ4Ł©ˇŁ
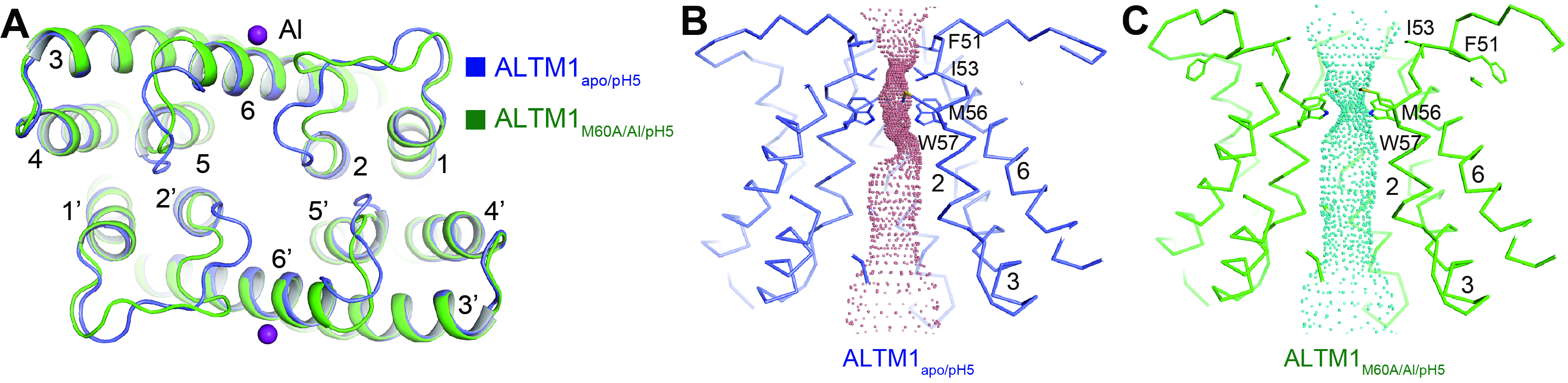
ÍĽ4 ϸ°űÍâ˛ŕÂÁŔë×ÓĽ¤»îAtALMT1ˇŁAŁ¬Î´˝áşĎÂÁŔë×ÓŁ¨Ŕ¶É«Ł©şÍ˝áşĎÂÁŔë×ÓŁ¨ÂĚÉ«Ł©µÄAtALMT1µÄ˝áąą±Č˝Ď¸©ĘÓÍĽˇŁBŁ¬Î´˝áşĎÂÁŔë×ÓµÄAtALMT1µÄŔë×Óͨ͸·ľ¶ˇŁCŁ¬˝áşĎÂÁŔë×ÓµÄAtALMT1µÄŔë×Óͨ͸·ľ¶ˇŁ
ÉĎĘöŃĐľż˝âÎöÁËAtALMT1¶ŕÖÖ״̬ϵÄČýά˝áąąŁ¬ÍęĂŔÚąĘÍÁËAtALMT1˝éµĽµÄÂÁĽ¤»îĆ»ąűËáתÔ˵ķÖ×Ó»úŔíŁ¨ÍĽ5Ł©Ł¬ÎŞ»ůÓÚµ°°×˝áąąÉčĽĆALMT1ą¦ÄÜ»ńµĂĐÔÍ»±äĚĺŇÔÔöÇż×÷Îď¸ůϵÂÁĽ¤»îµÄĆ»ąűËá·ÖĂÚŁ¬˝ř¶řĹŕÓýÄÍÂÁ×÷ÎďĚá¸ßËáĐÔÍÁČŔµÄÉú˛úÁ¦Ěáą©ŔíÂŰÖ¸µĽˇŁ
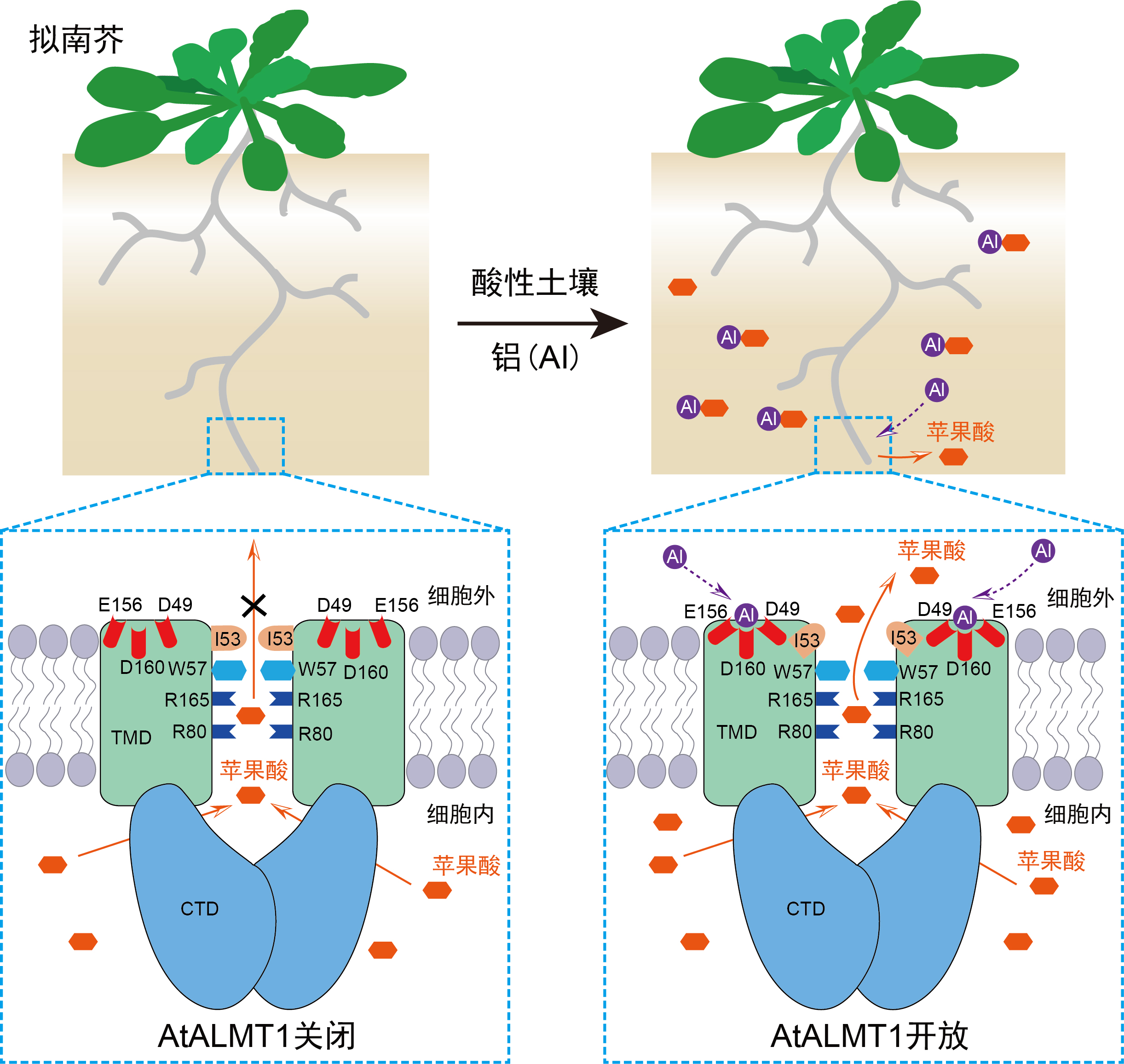
ÍĽ5 ÂÁĽ¤»îAtALMT1תÔËĆ»ąűËáµÄ·Ö×Ó»úŔíˇŁ
ąů˝ĚÎżÎĚâ×鲩ʿÉúÍő˝ÇÚˇ˘ŃîΡżÎĚâ×éÓŕĎķɲ©Ężˇ˘ÖŁÉÜ˝¨żÎĚâ×鶡ÖŇ˝ÜŃĐľżÔ±şÍÖĐżĆÔşÉîŰÚĎČ˝řĽĽĘőŃĐľżÔ±µÄŐĹСżµŃĐľżÔ±ÎŞÂŰÎĵŲͬµÚŇ»×÷ŐߡŁąů˝ĚΡ˘ŃîΡşÍÖŁÉÜ˝¨ÎŞą˛Í¬Í¨Ń¶×÷ŐߡŁ±ľŃĐľż»ąµĂµ˝ÁËÖĐąúşŁŃó´óѧĐěÎýĂ÷¸±ŃĐľżÔ±µÄÖ§łÖˇŁ±ľŃĐľżĘܿƼĽ˛żÖصăŃĐ·˘ĽĆ»®ˇ˘×ÔČ»żĆѧ»ů˝đÎŻˇ˘Őă˝Ęˇ˝ÜÇŕµČĎîÄżµÄ×ĘÖúˇŁ
ÔÎÄÁ´˝ÓŁşhttps://www.nature.com/articles/s41422-021-00587-6
˛ÎżĽÎÄĎףş
[1] Uexk¨ąll, H. & Mutert, E. Global extent, development and economic impact of acid soils. Plant Soil. 171, 1-15 (1995).
[2] Kochian, L. V. P., M. A. Liu, J. Magalhaes, J. V. Plant Adaptation to Acid Soils: The Molecular Basis for Crop Aluminum Resistance. Annu. Rev. Plant Biol. 66, 571-598 (2015).
[3] Sasaki, T., Yamamoto, Y. et al. A wheat gene encoding an aluminum-activated malate transporter. Plant J. 37, 645-653 (2004).
[4] Tashiro, M., Furihata, K., Fujimoto, T., Machinami, T. and Yoshimura, E. Characterization of the malate-aluminum (III) complex using 1H and 27Al NMR spectroscopy. Magn. Reson. Chem. 45, 518-521 (2007).