







近年来,随着生活方式改变和肥胖流行,2型糖尿病(T2D)已成为全球重大公共卫生问题。胰岛β细胞功能进行性衰竭是T2D发生发展的核心病理环节。胰岛内除了内分泌细胞外,还有包括巨噬细胞在内的多种免疫细胞,共同维持胰岛免疫微环境和功能稳态[1, 2]。在T2D进程中,胰岛炎症是β细胞功能损伤的重要病因[3-5]。近年来,大量临床研究提示,饮食和运动等生活方式干预是预防和治疗T2D的有效手段[6-8]。然而,生活方式干预对胰岛免疫微环境、β细胞身份和功能的影响,及其潜在调控机制有待阐明。
浙江大学基础医学院孟卓贤教授团队长期致力于胰岛和骨骼肌的代谢调控研究,先后揭示了染色质重塑因子BAF60a、BAF60c等在T2D 胰岛β细胞功能损伤中的关键作用和调控机制[9, 10]。在系统解析T2D胰岛β细胞功能由代偿到失代偿的多组学动态变化的基础上,构建了T2D的饮食干预小鼠模型,并在此基础上发现CTCF是饮食干预改善T2D胰岛β细胞功能的核心机制[8]。2026年 5月26日,浙江大学基础医学院孟卓贤教授团队联合浙江大学-爱丁堡大学联合学院陈迪教授团队以及南京医科大学第一附属医院付麒教授团队合作,在Life Metabolism杂志在线发表研究论文“Exercise preserves β-cell function in type 2 diabetes by reshaping intra-islet macrophage-β-cell crosstalk”(图1)。在课题组前期工作的基础上,建立了T2D 的早期运动干预小鼠模型,揭示了运动通过循环因子SPARC(Secreted protein acidic and rich in cysteine)介导的胰岛内巨噬细胞-β细胞互作重塑,改善β细胞和血糖稳态。浙江大学基础医学院博士生杨米棋、周燕萍、吴清倩(已毕业)为论文共同第一作者,孟卓贤教授、张哲雨博士、陈迪教授和付麒教授为共同通讯作者。

图1 论文于2026年5月26日在线发表
该研究通过高脂饮食(HFD)喂养结合跑台运动,建立T2D的早期运动干预小鼠模型(图2),发现运动显著改善T2D小鼠糖耐量和胰岛素敏感性,缓解HFD诱导的β细胞功能障碍、胰岛肥大及β细胞亚群失衡。同时,运动明显减少胰岛内CD45⁺免疫细胞浸润,抑制促炎基因表达,表明其对胰岛炎症具有显著抑制作用。通过非靶向血浆蛋白质组学,研究团队发现并验证循环中SPARC水平的下调可能是介导运动缓解T2D胰岛炎症和β细胞功能损伤的关键因子。机制研究显示,在高脂等应激条件下,SPARC通过激活巨噬细胞NLRP3炎症小体,促进Caspase-1依赖的IL-1β成熟释放,进而损伤β细胞功能;而运动介导的SPARC下调可阻断这一巨噬细胞-β细胞有害串扰。人群队列研究进一步证实,T2D患者循环SPARC水平显著升高,与胰岛素抵抗正相关而与β细胞功能和胰岛素敏感性相关指标呈显著负相关。
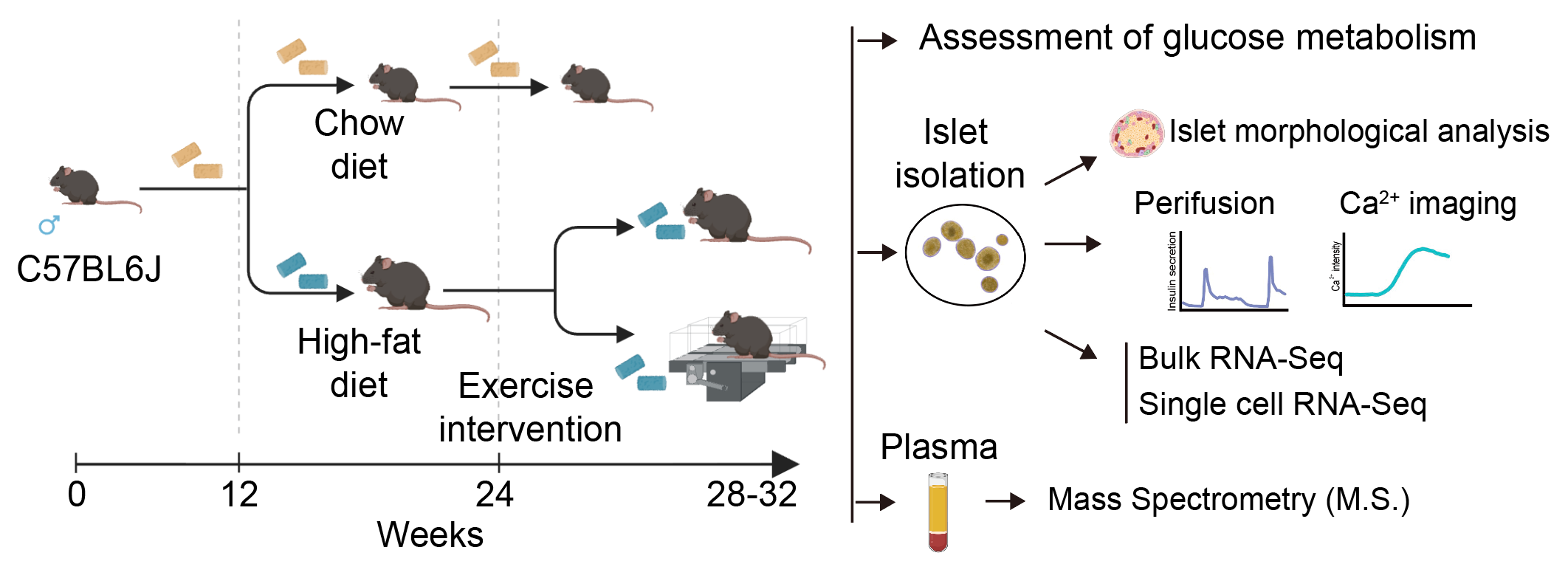
图2 运动干预小鼠造模示意图
为直接评估运动对胰岛形态和β细胞功能的影响,研究团队分离胰岛并进行全面检测。H&E染色结果显示,HFD诱导胰岛显著增生肥大,总胰岛质量增加4-5倍,而运动干预有效逆转了这一病理性改变,胰岛大小和数量恢复至接近正常水平。动态灌流实验进一步揭示,HFD会引发β细胞I相和II相葡萄糖刺激的胰岛素分泌(GSIS)功能严重受损,而早期运动干预几乎完全恢复了双相胰岛素分泌功能(图3)。

图3 运动保护胰岛形态及β细胞功能
已知胰岛内的β细胞具有显著异质性和可塑性。为进一步阐释运动干预对胰岛内β细胞亚群的影响,研究团队对运动模型小鼠的胰岛进行了单细胞转录组测序(scRNA-Seq)分析。结果显示,HFD引起保护型β1亚群(高表达Mt1、Mt2、Nupr1)比例显著减少,功能障碍型β2亚群(高表达Dapl1、Pdyn)比例显著升高,而运动干预能够几乎完全逆转上述变化,重塑β细胞组成和身份,改善β细胞功能(图4)。
图4 运动重塑T2D小鼠胰岛β细胞亚群
在阐明运动保护β细胞功能的同时,研究聚焦其对胰岛免疫微环境的影响。胰岛转录组测序结果证实,运动能够显著抑制HFD诱发的胰岛免疫活化,下调炎症相关基因表达,基因集富集分析结果亦与之相符。同时,胰岛流式细胞检测及免疫荧光染色实验发现,高脂饮食可使小鼠胰岛内CD45⁺免疫细胞比例增加近1倍,而运动干预可有效逆转该异常变化(图5)。
图5 运动改善T2D小鼠胰岛炎症及免疫细胞浸润
为寻找连接外周信号与胰岛保护的循环因子,研究团队富集了运动模型小鼠血浆低丰度蛋白并进行蛋白质组学分析,共鉴定出164个运动可逆调控的蛋白,其中SPARC在HFD中显著上调、运动干预后显著下调。组织表达谱显示,SPARC主要来源于脂肪组织和骨骼肌,HFD诱导其在多组织高表达,而运动则显著抑制这一变化(图6)。体外重组SPARC-Fc蛋白处理小鼠和人原代胰岛实验证实,SPARC明显抑制小鼠和人类原代胰岛的胰岛素分泌功能(图7)。
图6 发现循环因子SPARC可能介导运动改善胰岛功能
图7 SPARC 显著抑制人和小鼠胰岛的胰岛素分泌功能
机制研究表明,SPARC并不直接作用于胰岛β细胞,而是特异性结合并激活巨噬细胞。在巨噬细胞-β细胞共培养或条件培养基实验中,SPARC通过诱导巨噬细胞释放促炎因子显著损伤MIN6细胞GSIS。此外,使用Csf1r抗体清除胰岛驻留巨噬细胞后,SPARC对原代胰岛功能的抑制作用显著降低,同时β细胞亚群标志基因和胰岛素分泌基因表达得以恢复(图8)。进一步实验发现,在代谢应激条件下,SPARC特异性增强巨噬细胞NLRP3炎症小体激活,促进Caspase-1裂解和成熟IL-1β释放,而这一过程可被运动引起的SPARC下调所阻断。

图8 SPARC通过巨噬细胞损伤胰岛β细胞功能
最后,研究团队对包含99名糖耐量正常(NGT)个体和68名新诊断T2D(NDM)患者的人群样本进行了系统分析,结果显示NDM患者血浆SPARC水平显著升高,与胰岛素抵抗指标呈正相关,与胰岛素敏感性和β细胞功能相关指标显著负相关。这些结果为靶向开发SPARC作为T2D胰岛功能评估的生物标志物和干预靶点提供了临床证据(图9)。

图9 人群血液循环中SPARC水平与β细胞功能和胰岛素敏感性显著相关
综上所述,本研究揭示了运动通过下调循环因子SPARC,抑制胰岛巨噬细胞NLRP3-IL-1β轴重塑胰岛巨噬细胞-β细胞对话,从而保护2型糖尿病胰岛β细胞功能的全新分子机制(图10)。这一发现不仅系统解析了运动干预对胰岛免疫微环境、β细胞身份和功能的调控作用,更为开发针对SPARC-巨噬细胞-β细胞轴的精准干预策略提供了重要理论依据,有望为2型糖尿病的预防和治疗开辟新策略。
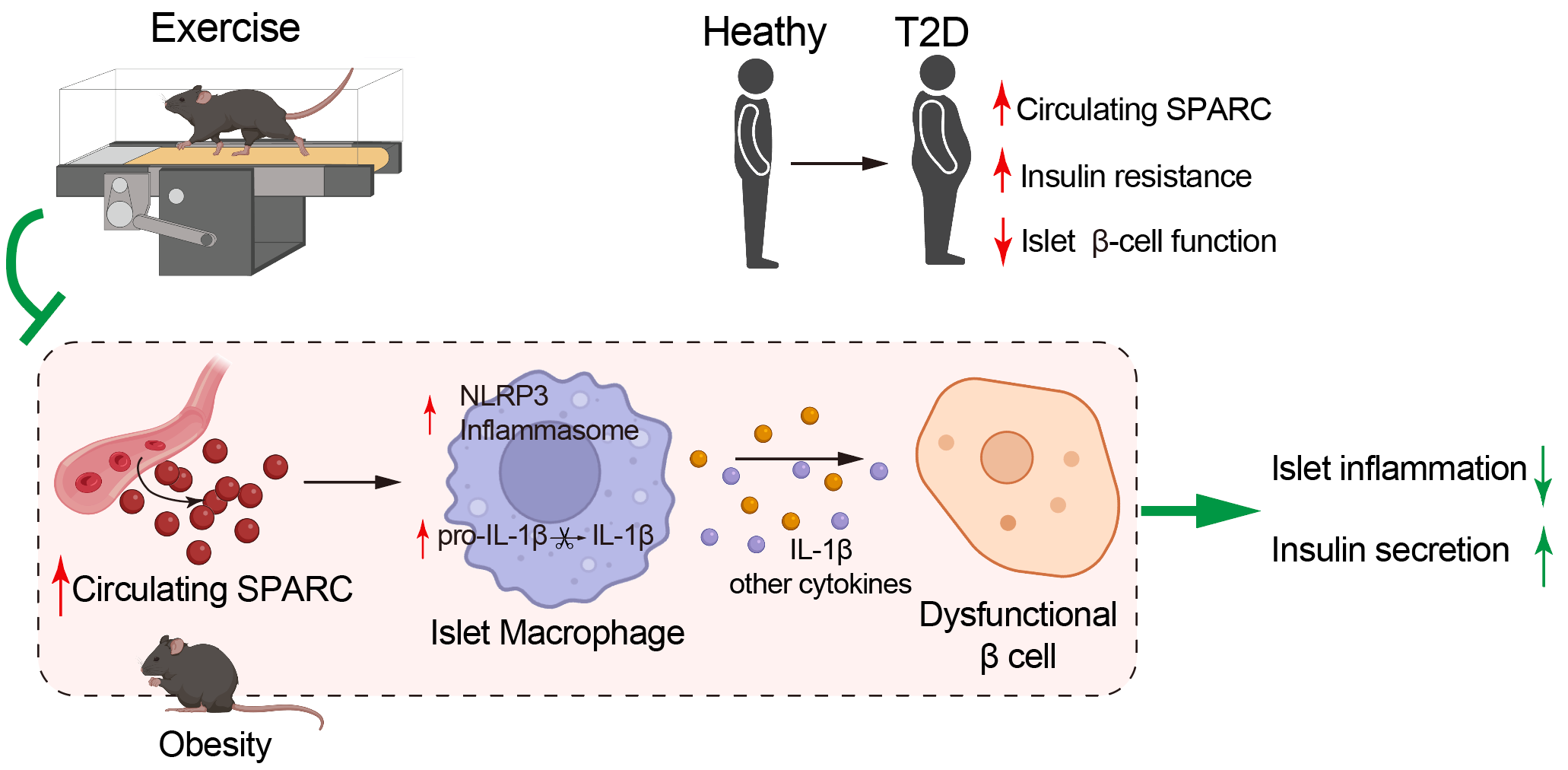
图10 运动通过下调SPARC重塑胰岛免疫微环境保护β细胞功能
1.Ying, W., et al. The role of macrophages in obesity-associated islet inflammation and beta-cell abnormalities. Nat Rev Endocrinol, 2020. 16(2): p. 81-90.
2.Grosjean, A., et al. An islet-resident macrophage antioxidant program preserves beta cell physiology. Sci Immunol, 2025. 10(113): p. eadz5181.
3.Kahn, S.E. Clinical review 135: The importance of beta-cell failure in the development and progression of type 2 diabetes. J Clin Endocrinol Metab, 2001. 86(9): p. 4047-58.
4.Cuenco, J. and Dalmas, E. Islet Inflammation and beta Cell Dysfunction in Type 2 Diabetes. Handb Exp Pharmacol, 2022. 274: p. 227-251.
5.Biden, T.J., et al. Lipotoxic endoplasmic reticulum stress, beta cell failure, and type 2 diabetes mellitus. Trends Endocrinol Metab, 2014. 25(8): p. 389-98.
6.Legaard, G.E., et al. Effects of different doses of exercise in adjunct to diet-induced weight loss on the AGE-RAGE axis in patients with short standing type 2 diabetes: Secondary analysis of the DOSE-EX multi-arm, parallel-group, randomised trial. Free Radic Biol Med, 2023. 208: p. 52-61.
7.Chen, Z., et al. Self-reported physical activity is associated with beta-cell function in Mexican American adults. Diabetes Care, 2013. 36(3): p. 638-44.
8.Wang, R.R., et al. Dietary intervention preserves beta cell function in mice through CTCF-mediated transcriptional reprogramming. J Exp Med, 2022. 219(7).
9.Qiu, X., et al. BAF60a-dependent chromatin remodeling preserves beta cell function and contributes to the therapeutic benefits of GLP-1R agonists. J Clin Invest, 2025. 135(23).
10.Yang, Z., et al. BAF60C links nucleolar stress to beta cell dysfunction in type 2 diabetes through controlling Reg3b mRNA decay. Dev Cell, 2026. 61(5): p. 976-994 e11.
